Development of Methods for Safe In Vitro-Differentiation of Different Progenitor Types in Osteoblasts/Osteoblast-Like and Osteoclasts/Osteoclast-Like Cells with Human and Mouse Origin
Sainova I.1*, Valkova I.2, Alexieva B.3, Nikolova E.4
DOI:
1* Iskra Sainova, Institute of Experimental Morphology, Pathology and Anthropology with Museum, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria.
2 Ilina Valkova, Institute of Experimental Morphology, Pathology and Anthropology with Museum, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria.
3 Bistra Alexieva, Institute of Experimental Morphology, Pathology and Anthropology with Museum, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria.
4 Elena Nikolova, Institute of Experimental Morphology, Pathology and Anthropology with Museum, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria.
Purpose: Development and application of techniques for safe derivation of cells from osteoblast and osteoclast lineages from progenitors of different sources and origin. Methods: Osteoblasts and osteoclasts with human origin were obtained from adult human bone marrow stem/stromal cells. Normal human mesenchymal stromal/stem cells (MSCs) were derived from human bone marrow material. Isolated nucleated cells were washed twice and resuspended in basic α-MEM (Minimum Essential Medium, alpha modification), supplemented with 15% heat-inactivated Fetal Bovine Serum (FBS), 50µg/ml freshly prepared Ascorbic acid (Vitamin C) and antibiotics (100µg/ml Penicillin G). Results: In both cases of human bone marrow material used, derivation of mature osteoclasts and osteoblasts was proved, when respective appropriate conditions/growth factors were present. Probably, separated stem/progenitor cells in general 3T3 line differentiated in each one of respective lineages, when appropriate cultivation conditions are available. Conclusions: Besides factors of cultivation and components of extra-cellular matrix, role of cryo-protector DMSO as a stimulator of both cell differentiation in two cell types, but also of intra-cellular fusion in formation of osteoclast-like cells, was suggested.
Keywords: Osteoblasts/Osteoblast-Like Cells Osteoclasts/Osteoclast-Like Cells Growth Factors/Cytokines Extracellular Matrix
| Corresponding Author | How to Cite this Article | To Browse |
|---|---|---|
| , , , Institute of Experimental Morphology, Pathology and Anthropology with Museum, Bulgarian Academy of Sciences, 1113 Sofia, , Bulgaria. Email:  |
Iskra Sainova, Ilina Valkova, Bistra Alexieva, Elena Nikolova, Development of Methods for Safe In Vitro-Differentiation of Different Progenitor Types in Osteoblasts / Osteoblast-Like and Osteoclasts / Osteoclast-Like Cells with Human and Mouse Origin. Biomed Rev J Basic Appl Med Sci. 2018;5(1):23-31. Available From http://www.biomedicalreview.in/progenitor-types-osteoblastsosteoblast-osteoclastsosteoclast-like-cells-origin-research-article |
 |
 ©
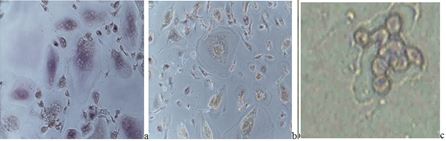
©  Fig. 1 In vitro-cultivated normal mouse embryonic 3T3 fibroblasts: a) control embryonic cell culture; b) mixed cell culture of 3T3 normal embryonic fibroblasts and malignant mouse myeloma cells – signs of early myeloid differentiation, as increased, rounded shape, but also light-stained cytoplasmic content with appearance of granules, centrally-located nuclei and changed nuclei/cytoplasm ratio, could be noted in comparison with the control fibroblasts (stained by Giemsa dye - magnification: x100)
Fig. 1 In vitro-cultivated normal mouse embryonic 3T3 fibroblasts: a) control embryonic cell culture; b) mixed cell culture of 3T3 normal embryonic fibroblasts and malignant mouse myeloma cells – signs of early myeloid differentiation, as increased, rounded shape, but also light-stained cytoplasmic content with appearance of granules, centrally-located nuclei and changed nuclei/cytoplasm ratio, could be noted in comparison with the control fibroblasts (stained by Giemsa dye - magnification: x100)